
EAP Publications | Virtual Library | Magazine Rack | Search | What's new
Ecological Solutions Roundtable
AGRO-BIO - 320 - 08
______________________________
Presque partout dans le monde, trois principaux lépidoptères peuvent causer des dommages importants aux cultures de crucifères. Il s'agit de la piéride du chou (imported cabbage worm - Pieris rapae), de la fausse arpenteuse (cabbage looper - Trichoplusia ni) et de la fausse-teigne des crucifères (Diamondback moth - Plutella maculipennis ou P. xylostella).
L'importance de l'un ou de l'autre de ces ravageurs varie avec les régions de la planète. Au Canada, dans la région d'Ottawa, Harcourt et al. (1955) a déterminé, sur une période de quatre ans, que la piéride causaient 85% des dommages, la fausse-teigne 13% et la fausse arpenteuse 2% dans les cultures de chou d'été. Dans les cultures tardives, la piéride causaient 66% des dommages, la fausse-teigne 26% et la fausse arpenteuse 8%. Comme il s'agit de papillons qui peuvent parcourir de grandes distances, il peut y avoir des années où ces proportions sont très différentes. Ainsi, dans le Sud-Ouest et le Nord de Montréal, la fausse-teigne représente depuis quelques années souvent un plus gros problème que la piéride, principalement en raison de la résistance aux insecticides conventionnels.
La piéride du chou (imported cabbage worm - Pieris rapae) occasionne des dommages importants dans les cultures de crucifères presque partout dans le monde. Au Québec, elle a été introduite accidentellement d'Europe en 1860. On l'a d'abord observé à Québec et, à peine quarante ans plus tard, on la retrouvait d'un océan à l'autre, de la Baie d'Hudson au Golfe du Mexique. Deux de ses proches cousines, la piéride des crucifères (Pieris napi) et la piéride occidentale du chou (Pieris occidentalis) causent des dommages le plus souvent négligeables en comparaison car elles préférent souvent les crucifères sauvages aux crucifères cultivées. Une autre espèce de piéride, que l'on ne retrouve heureusement pas ici, la grande piéride du chou (Pieris brassicae) est d'une importance économique dans les pays européens.
Les dommages sont causés par les larves qui se nourrissent du feuillage de plusieurs crucifères (voir tableau 1) mais plus particulièrement des Brassica (choux, choux-fleurs, choux de Bruxelles, brocolis). Les larves percent des trous circulaires sur les feuilles extérieures du chou et sur les premières feuilles de la pomme de chou. Elles pénètrent parfois jusqu'au coeur. Sur les choux-fleurs et brocolis, d'autres dommages résultent du dépôt par les larves d'excréments brunâtres ou verdâtres qui décolorent la tête. Ces dépôts se retrouvent aussi à l'angle des feuilles.
La piéride du chou est responsable de plus de la moitié des dommages au feuillage du chou. Dans l'est de l'Ontario, sur quatre ans, on a estimé que la piéride est responsable de 85% des dommages en culture hâtive de chou et 66% en culture tardive (Harcourt et al., 1955).
Comme il s'agit d'un insecte migrateur, les niveaux d'infestation dans une région donnée et d'une année à l'autre sont imprévisibles. Le niveau d'infestation dépend directement du nombre de papillons qui migrent et dans une moindre mesure, du nombre qui survivent à l'hibernation.
Les plantes attaquées par la piéride du chou sont surtout des crucifères cultivées ou sauvages mais aussi des plantes de trois autres familles peu ou pas représentées dans nos régions, soit des résédacées, des capparidacées et des tropaeolacées. Ces plantes ont toutes en commun des composés qui attirent la piéride appelés allyl-glucosinolates. Il s'agit de composés non volatiles à base de soufre(1). Ces composés, souvent toxiques pour d'autres insectes, agissent au contraire sur les piérides et autres papillons spécialistes des crucifères comme un incitatif à pondre et à s'alimenter.
Certaines crucifères n'ont pas de glucosinolates, ou ont d'autres composés qui les rendent peu attirantes et même répulsives à la piéride. C'est le cas de la bourse-à-pasteur (Capsella bursa-pastoris), une adventice assez courante, et de la vélar giroflée (Erysimum cheiranthoides).
Papillon
Le papillon mâle est d'un blanc glauque. Le bout de ses ailes avant est noir ou grisâtre et on dénote deux taches noires sur chaque paires d'ailes situées ainsi: une tache près du milieu de l'aile avant; une tache sur le bord supérieur de l'aile arrière.
La couleur de la femelle varie d'un blanc crémeux à un jaune terne. Elle possède aussi des extrémités d'ailes noires ou grises et des taches noires sur les ailes disposées de façon légérement différentes de celles du mâle. Les ailes avant ont chacunes deux taches noires et les ailes arrières une tache noire au bord supérieur. La grandeur moyenne des ailes une fois déployées est de 50 mm. La femelle est légérement plus grande que le mâle.
Oeufs
Les oeufs, en forme d'obus, sont à peine visibles à l'oeil nu (1 mm de long et 0,5 mm de largeur). Tout de suite après la ponte, ils sont d'un blanc pâle. Ils deviennent plus foncés pendant le développement de l'embryon pour finalement devenir jaune doré avant l'éclosion. Les oeufs sont attachés verticalement sous la feuille, étant attachés par leur base la plus large.
Larves
Les larves sont des chenilles de couleur vert feuille. Elles semblent être habillées de velours en raison de nombreux poils noirs et blancs qui forment une espèce de manteau transparent sur tout le corps. Elles sont marquées d'une fine bande orange ou jaune longitudinale sur le milieu du dos et d'une bande brisée de chaque côté. Elles possèdent trois paires de vrais pattes et cinq paires de fausses pattes charnues. Les chenilles, qui mesurent 3.2 mm à l'éclosion, atteignent 3 cm au dernier stade larvaire.
Chrysalide
La chrysalide, qui mesure 18 mm de longueur, est bombée au centre et pointue aux extrémités. Sa couleur est variable. Le plus souvent elle est brun pâle ou brun foncé, mais peut aussi être vert foncé ou blanche. Occasionellement, elle est vert pâle ou rosée. Comme c'est le cas pour de nombreuses espèces, la couleur peut varier de façon à ce que les chrysalides soient mieux camouflées dans leur entourage pour éviter la prédation. Cependant, les chrysalides de la piéride ont plutôt tendance à s'installer dans des endroits cachés, ce qui explique parfois leur piètre camouflage pour ce qui est de la couleur.
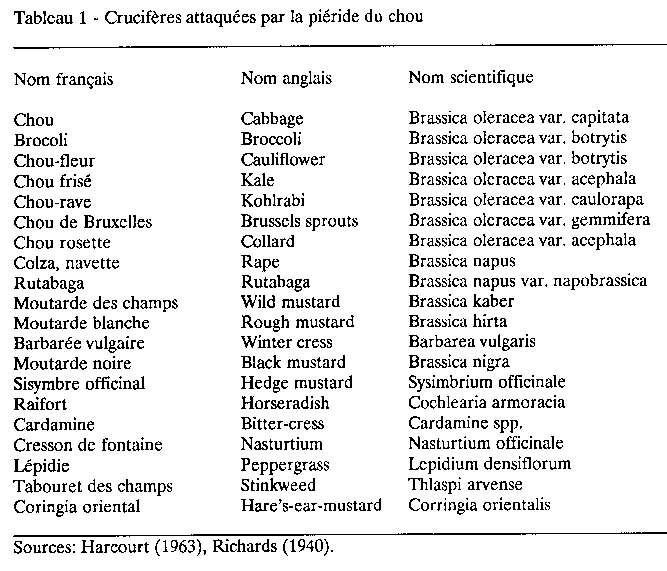
Au Québec, il y a généralement trois générations de piérides qui se chevauchent plus ou moins. La première génération passe souvent inaperçue tellement les populations sont faibles. Harcourt (1963) a étudié de près le cycle de vie de la piéride dans l'est de l'Ontario (région d'Ottawa). On peut considérer que le cycle de vie de l'insecte est à peu près le même dans une bonne partie du Sud du Québec. Le temps nécessaire pour passer de l'oeuf à l'adulte varie de 24 à 61 jours, étant en moyenne de 31 jours en juillet et août.
Stade adulte
Les premières piérides apparaissent de la mi-mai à la fin de mai. Ces premiers papillons proviennent surtout de chrysalides qui ont hiberné. Comme la survie à l'hiver n'est pas très élevée chez cette espèce dans nos climats, la première génération est en général moins abondante que les suivantes. Pour les deuxième et troisième générations, le vol migratoire en provenance de régions plus au sud jouent un rôle important dans l'abondance des populations (Harcourt, 1962). Il y a des épisodes de migration en mai, juillet et début d'août.
Les adultes vivent en moyenne une vingtaine de jours, soit l'équivalent de 156 à 220 degrés-jours au-dessus de 10C (Gossard et Jones, 1977). Le ratio de mâles et de femelles est à peu près égal bien que les mâles aient plus tendance à périr. L'accouplement a lieu en moins de 24 heures après la sortie du papillon du stade de chrysalide.
La ponte de la première génération se fait surtout dans les deux semaines suivant l'apparition des papillons, soit entre la mi-mai et la mi-juin. La plus grande période de ponte pour la deuxième génération dans l'est de l'Ontario a lieu dans la deuxième semaine de juillet et pour la troisième génération à la mi-août. Pendant sa courte vie d'une vingtaine de jours, la femelle peut pondre jusqu'à 400 oeufs.
Oeufs
Les oeufs éclosent en moins d'une semaine par temps chaud. La période d'incubation dans l'est de l'Ontario varie de 4 à 8 jours, 5 en moyenne en juillet et août.
Larves
Les larves peuvent se développer entre 7 et 30C, des températures supérieures à 12C étant préférables. Les larves, qui subissent quatre mues, passent donc par cinq stades. Elles sont le plus vorace à des températures moyennes (19C). A des températures hautes (24C) ou basses (<13C) les dommages diminuent (Tatchell, 1981). La période larvaire varie de 12 à 33 jours ou plus, 15 en moyenne en juillet et août. Au Québec, on peut observer des chenilles de juin à octobre avec des pointes de la mi-juillet à la fin d'août.

Pendant sa croissance la chenille se déplace très peu et reste en général sur le même plant. Elle demeure même sous la feuille extérieure qui l'a vu naître pendant les trois premiers instars. Elle se déplace ensuite vers le centre de la plante et s'attaque voracement aux parties commercialisables de la plante. C'est donc les deux derniers instars qui causent la majorité des dommages. Après son développement complet, la chenille repose et erre pendant un temps variable (environ 2 jours) et passe au stade de chrysalide.
Nymphose
La durée de la nymphose varie de 8 à 20 jours, en moyenne 11 jours en juillet et août. La nymphose ne se fait sur la plante-hôte que pour la première génération de chenilles. Les sites de nymphose pour les autres générations sont variés et semblent dépendre de la densité de population. En certains cas, la nymphose a lieu sur le plant même ou dans les environs immédiats. En d'autres cas, les chenilles font des distances considérables et s'installent sur des poteaux de clôtures, des bâtiments, etc.
Caractéristiques générales de vol
Le vol des piérides est lent et irrégulier sauf si elles sont emportées par le vent. Elles ont en fait tendance à voler contre le vent. Leur vol s'oriente souvent vers le nord, surtout par haute pression quand il y a des vents chauds du sud.
Les piérides volent toute la journée sauf si les conditions sont défavorables. Les conditions les plus favorables au vol, à l'accouplement et à la ponte sont des hautes températures, des vents faibles et un grand ensoleillement. Au contraire, le froid, la pluie et les grands vents limitent leurs activités.
Les femelles gravides sont actives pendant tout le jour, volant une moyenne de 11 heures par jour. Elles commencent leur journée deux heures après le levée du jour et sont le plus actives vers midi.
Ponte
La piéride dépose ses oeufs un à la fois presque exclusivement sous la feuille. Sur les plantes à feuilles larges, elle se tient sur le côté d'une feuille horizontale et dépose son oeuf très près du bord de la feuille. Pour éviter d'être emporter par le vent, elle traverse un champ en faisant face au vent et pond du côté vers lequel le vent souffle.
Taux de ponte
Le fait que les piérides ne pondent qu'un oeuf à la fois ne veut pas dire qu'elles pondent un seul oeuf par plant. Selon des observations dans l'état de New York (Root et Kareiva, 1984), elles peuvent pondre jusqu'à 80 oeufs par plant en certains cas. Beirne (1971) rapporte que dans les champs non-traités de la région d'Ottawa, on observe en moyenne 5,8 chenilles par plant par saison.
La variation d'année en année du nombre d'oeufs sur les cultures dépend de l'abondance des papillons mais aussi de l'attirance des lieux pour ceux-ci. Quoi qu'il en soit, les conditions climatiques sont déterminantes. Les femelles peuvent ainsi ne pas pondre à leur plein potentiel en raison du mauvais temps. Un épisode de gel réduit drastiquement la ponte.
Gossard et Jones (1977) ont étudié le taux de ponte en fonction des données climatiques. Ils ont établi que:
Choix du site de ponte
Il s'est fait beaucoup de recherches sur le comportement des piérides quant à la sélection d'un site de ponte. Le principal élément à retenir est que, contrairement à beaucoup d'insectes qui ont tendance à être plus abondants dans une monoculture, la piéride a tendance à être moins abondante dans un grand champ ou une zone dense de crucifères que sur des plantes-hôtes dispersées à travers d'autres plantes diverses. Ce comportement s'explique par les besoins alimentaires des papillons mais constitue aussi à une façon d'augmenter les chances de survie des chenilles. En effet, comme les jeunes chenilles sont peu mobiles, il est préférable pour les femelles piérides de ne pas pondre sur les plants qu'elles ont déjà visités afin d'augmenter les chances de survie des larves.
Le choix du site de ponte dépend de facteurs visuels, tactiles et olfactifs. Les facteurs qui ont fait l'objet de recherches sont: la grosseur et l'emplacement des parcelles; le couvert végétal; la proximité de sources de nourriture ou d'autres plantes-hôtes; la composition ou qualité de la plante-hôte; l'aspect extérieur de la plante-hôte; et la présence de phéromones de ponte.
Grosseur et emplacement des parcelles: Cromartie (1975a et b) a étudié l'effet de la grosseur et de l'emplacement des parcelles de crucifères sur l'attirance de la piéride. En comparant des parcelles de 1, 10 ou 100 choux rosettes, il a établi que plus les parcelles de crucifères étaient grosses, moins la piéride causait de dommages en proportion. Il a aussi observé que les femelles pondent très peu souvent dans les bois ou sur les bords de boisé où pourtant poussent des crucifères. Elles préfèrent les endroits ouverts et ensoleillés.
Couvert végétal: La piéride a tendance à être plus abondante lorsque le couvert végétal est varié. Plus de détails à ce sujet sont donnés dans la section sur la diversité végétale dans les moyens de lutte culturaux.
Proximité des sources de nourriture: Les piérides pendant la journée alternent entre les plantes où les larves se développeront et les fleurs sauvages où elles peuvent s'alimenter. On observe donc en général plus d'oeufs sur les plantes en bordure des champs qu'au centre parce que les femelles préfèrent être près de leur source de nourriture.
Qualité de la plante-hôte: L'âge et la composition de la plante et son contenu en eau sont tous des facteurs qui peuvent agir sur l'attirance de la piéride. La piéride est très efficace à repérer les crucifères mais n'est pas très bonne à repérer les plants déjà visités ou à évaluer la qualité des plantes-hôtes selon Root et Kareiva (1984). Dans leurs expériences, ils ont observé les piérides déposant des oeufs sur des plants de chou rosette fanés et contenant très peu d'azote, et sur des plants où de nombreux oeufs avaient déjà été déposés. Plus de détails sur ce sujet sont donnés dans la section sur la fertilisation dans les moyens de lutte culturaux..
Aspect extérieur des plants: Renwick et Radke (1988) ont établi que la grandeur et la forme des plantes avait peu d'importance dans leur sélection par la piéride. La couleur était le plus important facteur avec la présence de glucosinolates et autres composés propres aux crucifères.
Présence de phéromones: En Europe, la grande piéride Pieris brassicae produit une phéromone qui lui sert à indiquer les plantes où elle ou ses consoeurs ont déjà pondu. Elle peut ainsi éviter de pondre sur ces plantes et donc augmenter les chances de survie des larves. Or, la piéride du chou perçoit également cette phéromone. En appliquant cette phéromone sur des plants, Klijnstra (1985) a pu empêcher la ponte de P. rapae pendant plus de sept jours.
Plus récemment, Schoonhoven et al. (1990) ont démontré l'existence d'une phéromone semblable chez la piéride du chou en couvrant les feuiles de chou avec une solution d'oeufs de piéride broyés. Cette phéromone le plus souvent n'agit pas en contexte naturel en raison de différents facteurs dont la disponibilité et la répartition des sites de ponte et la qualité des plantes.
Distribution de la ponte
L'observation du comportement de ponte révèle que les femelles pondent le plus souvent de façon irrégulière. Schoonhoven et al. (1990) ont dénoté que la distribution de la ponte change selon la densité de population. Quand la population de papillons est faible, les piérides vont souvent concentrer leurs efforts sur une petite section, voler quelques temps en délaissant de grands secteurs et puis de nouveau concentrer leurs efforts sur une autre section. Au contraire, quand la population est forte, les femelles ont tendance à pondre plus régulièrement et à bien couvrir tout le territoire.
Avant d'aborder les moyens de lutte, il est important de signaler qu'une espèce prolifique comme la piéride du chou est sujette à une grande mortalité naturelle, sans quoi les populations atteindraient vite des niveaux catastrophiques. Ainsi, en plus d'un faible taux de survie des chrysalides à l'hiver, les adultes meurent parfois lorsqu'au printemps leur vol migratoire les amène dans une zone où les plantes dont ils se nourrissent ne sont pas disponibles. Les larves, surtout les plus jeunes, peuvent mourir par noyade lorsque de fortes pluies créent des accumulations d'eau à la base des feuilles ou les délogent et les font tomber dans des flaques d'eau. Tous les stades de l'insecte, et les larves particulièrement, succombent à plusieurs maladies et prédateurs qui seront abordés dans la section sur les moyens de lutte biologiques.
Dans le cas des lépidoptères ravageurs, il est recommandé d'effectuer le dépistage pour chaque champ de Brassica et non pas seulement dans un champ représentatif (Andaloro et al., 1982). Au Québec, les dépisteurs examinent la totalité du plant car ils recherchent en même temps les chenilles de fausse-teigne et de fausse arpenteuse qui se tiennent dans des endroits différents sur le plant.
Il est recommandé de vous adresser aux bureaux locaux ou régionaux du MAPAQ pour connaître le nom de technologistes ou d'agronomes qui font du dépistage dans les cultures de crucifères dans votre région. Le Manuel de l'observateur - légumes disponible par le MAPAQ est également un document utile pour le dépistage.
Seuils de tolérance
Les seuils de tolérance à considérer varient selon la culture, le stade de la plante et le marché auquel elle est destinée. Ainsi, pour des choux destinés au marché frais, le seuil économique considéré par Kirby et Slosser (1981) était aussi bas que 0.30 larve/plant. Pour des choux destinés à la transformation (choucroute, salade de chou), le seuil peut évidemment être plus élevé. Sur des choux fraîchement transplantés, Samson et Grier (1983) ont établi que 7 larves se nourrissant pendant neuf jours ne réduisaient pas les rendements alors que 34 larves pendant 20 jours diminuaient le rendement de 34%. Les seuils acceptables en production biologique sont encore à établir.
Piégeage
Le piégeage des piérides n'est pas un sujet très développé. Hovanitz et Chang (1964) ont étudié l'effet de la couleur et de la concentration de glucosinolates sur l'attirance de la piéride. Ils ont observé que, en ordre décroissant, le bleu-vert, le vert, le vert-jaune et le bleu l'attirait le plus. Le blanc était peu attirant, le jaune était répulsif tandis que le rouge ne l'attirait pas du tout. Du côté olfactif, l'extrait de graines de moutarde à 2% est la concentration la plus attirante, les concentrations supérieures étant même répulsives selon les mêmes chercheurs. Des plants de brocolis feraient sans doute d'excellents pièges.
Comme mentionné précédemment, la qualité des crucifères, leur composition, peut avoir un effet sur l'attirance des femelles. Comme la fertilisation joue un rôle au niveau de la concentration des composés aromatiques et de la teneur en eau des plants, elle peut donc avoir une influence sur l'attirance de la piéride. Plusieurs chercheurs se sont penchés sur ce sujet. L'influence de la fertilisation était plus marquée en laboratoire qu'en champ.
Azote: Slansky et Feeny (1977) ont déterminé en laboratoire que le contenu en azote du feuillage affecte directement le développement des larves de piérides. Plus le feuillage contient d'azote, plus la larve se développe rapidement. Par contre, les larves sur plants avec peu d'azote ont tendance à consommer plus de feuillage et à mieux utiliser l'azote, de tel sorte que le contenu en azote des plants affecte finalement peu le taux de gain des larves.
Bien qu'en laboratoire les piérides démontrent une nette attirance à pondre sur les plants ayant reçus une fertilisation azotée abondante, en champ la même tendance n'a pas toujours été démontrée selon Létourneau (1988). Dans des expériences en champ faites en Floride, Jansson et al. (1991) ont démontré que lorsqu'on augmente les niveaux de fertilisation azotée, on accroît du même coup les problèmes de lépidoptères dans les cultures de choux. Ils ont comparé quatre niveaux de fertilisation, soit 0, 45, 90 et 168 kg/ha, le dernier niveau correspondant aux recommandations officielles en Floride. La relation entre le niveau d'azote et les populations de la piéride n'étaient toutefois pas aussi nette qu'avec les autres lépidoptères.
Azote et soufre: Comme les glucosinolates sont des composés à base de soufre, il est intéressant de voir l'effet de la fertilisation soufrée sur l'attirance de la piéride. Wolfson (1980) a comparé en chambre de croissance l'effet de quatre niveaux de fertilisation azotée et soufrée sur l'attirance de la piéride pour la ponte, en faisant la relation avec la concentration en eau et en composés soufrés de la moutarde noire. Elle a conclu que:
Azote et phosphore: Myers (1985) a établi que les femelles préféraient les plants ayant des plus grandes concentrations d'azote et de phosphore, un plus haut taux de transpiration et une couleur verte plus prononcée. Les femelles pouvaient détecter la différence seulement un jour après la fertilisation! La différence dans le taux de ponte était encore plus marquée après cinq jours alors que les plants devenaient plus vert. Les plants fertilisés avec des engrais très solubles auraient donc pour effet de rendre les plants plus attirants aux piérides.
Fertilisation organique vs inorganique: Eigenbrode et Pimentel (1988) ont examiné l'effet d'une fertilisation organique à base de fumier de vache sur les populations de ravageurs dans une culture de chou-rosette dans l'état de New York. Leurs traitements consistaient en: 220 t/ha de fumier frais appliquées au printemps; 220 t/ha appliquées en surface à l'automne; 12 t/ha frais appliqué au printemps; 785 kg/ha de 13-13-13; et un témoin non fertilisé. Ils ont observé que:
La diversité végétale a un effet certain sur l'équilibre des populations d'insectes. Dans les cultures, la présence de plusieurs espèces végétales sur le même sol se fait naturellement par la croissance des mauvaises herbes. D'autres façons d'accroître la diversité végétale sont d'implanter des plantes couvre-sols, des cultures intercalaires ou de pratiquer le compagnonage.
Adventices: Les mauvaises herbes, indésirables du point de vue des rendements, ont un effet variable et plutôt neutre sur la piéride. L'attirance des piérides peut être plus ou moins grande dûe à la présence de mauvaises herbes alors que la mortalité naturelle des larves augmente grâce à ces dernières. Dans l'état de New York, Cromartie (1975a) observait que la piéride était le seul ravageur des crucifères à envahir les parcelles où les mauvaises herbes n'étaient pas contrôlées. Smith (1976) en Angleterre a toutefois noté une plus grande population de larves et de chrysalides là où des choux de Bruxelles étaient sarclés (la couleur du sol semblait jouer un rôle dans ce cas). Dans une culture de choux de Bruxelles en Angleterre, Dempster (1969) a observé que la mortalité des larves de piérides était près du double (environ 70%) là où les mauvaises herbes n'étaient pas contrôlées. Il attribuait cette différence surtout à l'action des prédateurs, car il n'y avait pas plus de mortalité dûe aux maladies ou aux parasites. Yepsen (1984) suggère d'enlever les crucifères sauvages qui pourraient croître dans les champs et jardins pour éviter d'attirer les piérides. Comme les Brassica sont les plantes préférées des piérides, il semble douteux qu'une telle mesure puisse réduire appréciablement les populations.
Des chercheurs des Pays-Bas ont obtenu une réduction significative de tous les ravageurs des choux de Bruxelles à l'exception de la piéride en intercalant de la spargoute des champs (Theunissen et Den Ouden, 1980). La spargoute, ici considérée uniquement comme une mauvaise herbe, est parfois utilisée comme fourrage en Europe.
Couvre-sol de trèfle: Dempster et Coaker (1974) ont vérifié si un couvre-sol de trèfle blanc pouvait accroître la prédation des larves de piérides sans trop nuire à une culture de choux-fleurs. Le trèfle implanté à pleine superficie peu avant la transplantation des choux augmentait la prédation mais réduisait les rendements de façon importante. Dans le Midwest américain, Andow et al. (1986) ont étudié l'influence d'un couvre-sol permanent de trèfle blanc sur les populations d'insectes dans une culture de chou. Ils ont observé que la première génération de piéride était plus abondante avec le couvre-sol de trèfle que sans couvre-sol ou qu'avec des couvre-sol de pâturin et de fétuque. Par contre, la deuxième génération était moins abondante avec le couvre-sol de trèfle. L'utilisation d'un couvre-sol permanent en plus d'avoir un effet incertain sur les ravageurs réduisait trop les rendements de chou pour s'avérer une solution à envisager.
Ces expériences semblent indiquer que si un couvre-sol de trèfle est considéré, il ne doit pas être permanent, mais plutôt implanté en cours de saison. Un couvre-sol de trèfle rouge nain, par exemple, qui serait semé après un premier ou deuxième sarclage permettrait d'encourager les prédateurs de la piéride sans trop nuire à la culture.
Plantes nectarifères: Au Michigan, Zhao et al. (1992) ont observé un accroissement du nombre de larves et d'oeufs de piéride dans un champ où des plantes nectarifères avait été complantées avec du brocoli. En effet, les plantes nectarifères ont fourni l'environnement idéale de ponte pour les femelles qui n'avaient plus à aller au bord du champ pour se nourrir.
Compagnonage: Le compagnonage, c'est-à-dire l'association de deux ou plusieurs cultures, est souvent employé à petite échelle pour prévenir les problèmes de ravageurs. Les plantes compagnes recommandées par les manuels de jardinage biologique pour repousser la piéride sont le romarin, le thym et la sauge. Le céleri, la cathaire, la menthe et l'absinthe sont aussi mentionnés. Yepsen (1984) rajoute l'oignon, l'ail, la tomate, la tanaisie, le chanvre et l'hysope. Malheureusement, dans le cas de la piéride, les recherches scientifiques indiquent que le compagnonage n'est pas utile et même déconseillable que ce soit avec des plantes aromatiques ou d'autres légumes.
Plantes aromatiques: Comme les piérides sont attirées par l'odeur des crucifères, il est en théorie possible que la présence de plantes à fortes odeurs arrivent à masquer l'odeur des crucifères et donc à berner la piéride. En Virginie, Latheef et Irwin (1979) ont étudié l'effet de six plantes compagnes sur les lépidoptères s'attaquant aux crucifères. Aucune des plantes étudiées (tagètes, cresson, menthe poivrée, sauge, thym et hédéoma) n'a affecté positivement ou négativement les dommages dûs à la piéride. Toutes avaient tendance à compétititioner le chou-rosette, la tanaisie étant franchement allélopathique. Courter et Randell (1975) n'ont pas non plus observé de différences dans le nombre de larves de piérides lorsque des choux étaient complantés avec du thym ou de l'hysope. Les seuls résultats positifs (diminution significative du nombre de larves) de compagnonage avec des plantes aromatiques contre la piéride ont été rapportés par Koehler et al. (1983). Cela consistait à planter quatre plants d'anis pour un plant de chou (pas très pratique à moins de faire la culture de l'anis!).
Tomate: Il s'est fait beaucoup de recherche sur le compagnonage des Brassica et des tomates. Dès 1915, Vostrikov notait qu'une pratique générale dans une certaine région de la Russie consistant à intercaler des tomates et des choux permettait de diminuer les dommages de la piéride et de la fausse-teigne de façon importante. Par la suite, plusieurs études ont démontré que le compagnonage avec la tomate permettait de diminuer les populations de ravageurs des crucifères en général. En Nouvelle-Écosse, l'entomologiste Charles Fox d'Agriculture Canada cité par Bennett et Forsyth (1984) a obtenu 42% plus de dommages des ravageurs des crucifères dans une monoculture en comparaison avec une polyculture de deux rangs de choux pour trois rangs de tomates. En Inde, Srinivasan et Veeresh (1986) ont étudié l'effet de différentes façons de complanter les tomates et les choux sur les lépidoptères attaquant les choux. La façon qui réduisait le plus les dommages des lépidoptères consistait à complanter une rangée de choux et une rangée de tomates mais de transplanter les choux 30 jours après les tomates, ce qui affectait grandement les rendements de choux.
Malgré qu'il semble envisageable dans le cas des autres ravageurs des crucifères, le compagnonage avec la tomate ne réduit toutefois pas les populations de piérides. Il peut même avoir l'effet contraire à certains moments. Maguire (1984) a ainsi noté dans ses expériences un acroissement du nombre de larves sur des choux rosette complantés de plants de tomate cerise. Elle attribuait notamment ce phénomène à l'attirance des fleurs jaunes de la tomate alors qu'il n'y avait plus de crucifères sauvages (fleurs jaunes également) de disponibles pour les piérides.
On peut conclure qu'il n'y a pas d'intérêt à utiliser des plantes compagnes ou cultures intercalaires pour lutter contre la piéride. Dans certains cas, il peut toutefois être intéressant de tabler sur la diversité végétale pour contrôler d'autres ravageurs comme la mouche du chou. Il faut aussi noter que beaucoup des plantes mentionnées précédemment peuvent être d'excellents répulsifs (voir la section sur les répulsifs végétaux) qui décourageront la ponte de la piéride lorsque pulvérisés sur le feuillage des crucifères.
Différences de résistance entre les Brassica: Harrison et Brubaker (1943) ont évalué les différences de résistance entre les différentes types de Brassica. Ils ont observé plus de larves sur le chou rouge que sur le chou vert, tandis que le brocoli et le chou rosette étaient les plus attaquées. Ils concluent toutefois de leurs expériences que la variation du nombre de larves dépend avant tout d'autres facteurs que le type de Brassica, entre autres la quantité de feuillage produite par le plant, son stade de maturité et sa condition générale.
Radcliffe et Chapman (1966) ont observé que le chou chinois et le navet sont peu attaqués en raison des pilosités sur leur feuillage qui découragent la ponte. La moutarde attire de nombreux papillons, mais les femelles s'abstiennent généralement d'y pondre.
Dans une comparaison de la résistance de différents Brassica à la piéride, Pimentel (1961) a observé un peu moins de larves sur le chou rosette et le chou frisé que sur le brocoli, le chou de Bruxelles et le chou. Pimentel considère que la résistance est dûe non seulement à des facteurs internes (physiologie de la plante) mais aussi à des facteurs extérieurs. Il attribuait le moins grand nombre de chenilles sur le chou frisé et le chou rosette à un plus grand parasitisme parce que les feuilles de ces plantes exposaient plus les larves et les oeufs aux parasites que les autres crucifères.
Choux: Brett et Sullivan (1974) ont étudié la résistance à la piéride de dizaines de variétés de choux en Caroline du Nord. Ils ont observé que la résistance était semblable pour la fausse arpenteuse et la piéride. Les variétés qui se sont montrés très résistantes pendant 6 années sont: Mammoth Red Rock, Chieftan Savoy et Savoy Perfection Drumhead. Les variétés les plus susceptibles furent Golden Acre, Elite Golden Acre et Copenhagen Market 86.
Le chou rouge, peu importe son bagage génétique, est moins attirant pour la piéride que le chou vert en raison de sa couleur (Radcliffe et Chapman, 1966). De plus, le chou rouge est plus résistant que le chou vert à la première génération de piérides, mais sa susceptibilité s'accroît avec l'avance de la saison (Radcliffe et Chapman, 1965). Le changement de résistance avec la saison dépend du niveau de dommages précédents. Ainsi, les femelles piérides peuvent détecter les plants endommagés et décider de ne pas pondre sur ceux-ci afin d'augmenter les chances de survie de leurs larves. Comme les choux rouges sont moins attirants que les verts quand ils sont petits, ils sont épargnés en début de saison alors que plus tard les piérides seront attirés par le fait qu'ils n'ont pas été endommagés.
Brocoli: Vail et al. (1991) ont étudié la résistance de 13 variétés de brocoli à la piéride et autres lépidoptères en Virginie. Ils ont observé le moins grand nombre de larves sur les variétés hâtives et le plus grand nombre sur les variétés tardives. Ainsi, le cultivar Symphony avait le moins grand nombre de larves. Il n'y avait toutefois pas de différences au point de vue de la défoliation ou du pourcentage de brocoli commercialisable.
La rotation est appropriée à l'echelle d'un jardin et de toutes façons essentielle en champ pour éviter les problèmes de maladies des crucifères. Le nettoyage après la récolte d'une culture hâtive est important. Les résidus laissés au champ peuvent attirer les lépidoptères et faire en sorte que ce champ serve de foyer d'infestation dans les années à venir.
Comme les crucifères sont des plantes de climat frais, on peut éviter de cultiver des Brassica, particulièrement les plus sensibles, dans les périodes de pointe de la piéride (juillet et août) pour réduire de beaucoup le niveau de dommages.
D'autres problèmes que les ravageurs peuvent alors se poser. Ainsi, dans les cultures hâtives, il peut être difficile pour les crucifères d'obtenir l'azote nécessaire avec une fertilisation organique uniquement. De plus, la protection des jeunes plants contre le gel sera nécessaire. Dans les cultures tardives, le gel peut aussi poser des problèmes mais la période nécessaire à la maturation sera plus critique.
Couvertures
La protection physique la plus courante est la toile protectrice de polyéthylène, de nylon ou de coton fromage. Ces couvertures flottantes, qui servent à empêcher les femelles de pondre sur les plants, sont particulièrement indiquées pour les jeunes plants de Brassica partis à l'été, alors que les populations de papillons sont au plus fort. Les toiles ont aussi l'avantage d'accélérer la maturité des Brassica et de les protéger des gels légers en début de saison. Elles sont disponibles en différentes largeurs et longueurs (jusqu'à 15 par 250 mètres).
La couleur blanche des toiles est déjà peu attirantes pour la piéride mais la texture à aussi un rôle à jouer. Hovanitz et Chang (1964) ont testé différentes textures de couvertures en laboratoires pour savoir lesquelles repoussait le plus la piéride. Le papier blanc s'est avéré le moins attirant suivi du papier ciré.
Eau
Il est possible de déloger les larves en dirigeant un jet d'eau puissant sur les plants. Les larves, surtout les plus jeunes, ont rarement la force de remonter sur les plants et seront peut-être noyées au pied des plants. Contre la fausse arpenteuse, Talekar et al. (1986) ont utilisé avec succès l'irrigation au fusil (sprinkler) pendant cinq minutes au-dessus des plants tous les soirs au crépuscule. L'effet était de noyer les larves et de perturber le vol et la ponte des adultes. Avec cette méthode, il est important de faire attention à la structure du sol et de ne pas aggraver les problèmes de maladie.
La plupart des rapports du début du siècle indiquaient que la piéride était contrôlée de façon satisfaisante (pour l'époque!) par des insectes parasites et surtout par des maladies virales. Les plus importants facteurs de mortalité dûs à des agents biologiques sont en ordre décroissant: les maladies virales (environ 60%), les guêpes parasitaires (environ 30%) et les oiseaux (moins de 10%).
L'action des prédateurs est considérée comme peu importante dans le contrôle de la piéride, mais serait plus apparente chez les crucifères sauvages que chez les cultivées. Quoiqu'il en soit, il est bon d'encourager la présence des prédateurs pour l'équilibre de la ferme.
Punaises: L'hémiptère Reduviolus lativentris est un prédateur des oeufs de la piéride. La punaise pentatome Euschistus servus euschistoides, commune dans nos régions, se nourrit de larves de piérides selon Culliney (1985) mais de façon non spécialisée comme beaucoup de punaises qui se nourrissent aussi bien d'insectes que de plantes. Les punaises anthocores peuvent s'attaquer à tous les stades de l'insecte. Il existe sur le marché américain des produits qui permettent d'attirer les punaises prédatrices du genre Podisus, des prédateurs actifs de plusieurs espèces de chenilles.
Guêpes prédatrices: Les guêpes prédatrices Polistes et Vespula germanica sont des prédateurs redoutables des larves lorsque leurs populations sont suffisantes. Au Wisconsin, Gould et Jeanne (1984) ont établi des colonies de guêpes prédatrices Polistes dans des parcelles de choux en leur fournissant des nids artificiels constitués de boîtes de bois de 15x11x14 cm avec un fond de grillage de 1 cm et placés à 50 cm du sol. L'avantage de ces guêpes est qu'elles s'attaquent même aux très grosses larves de piérides. Un nid abritant 5.6 guêpes en moyenne était suffisant pour réduire de 44% le nombre de larves par chou par parcelle de 20 choux. Un taux encore plus élevé de prédation aurait pu être obtenu si les guêpes n'avaient pas eu accès à un champ de luzerne voisin où il existait de nombreuses autres proies. Dans les climats froids comme le nôtre, on doit établir la colonie en trouvant une colonie hivernante et en la transportant dans le lieu de culture.
Fourmis: Les fourmis sont des prédateurs des oeufs. En Australie, les fourmis sont considérées comme des prédateurs importants des larves (Jones, 1987).
Oiseaux: En Ontario, selon Harcourt (1962), les oiseaux qui ont été observés le plus souvent à manger les larves sont le vacher à tête brune (Molothrus ater), le moineau chanteur (Melospiza melodia) et le carouge à épaulettes (Agelaius phoenicus). En général, les oiseaux n'aiment pas trop les papillons adultes en raison de substances toxiques qu'ils contiennent. En Angleterre, Baker (1970) a observé le moineau (Passer domesticus) et la fauvette (Sylvia borin) se nourrir des oeufs de piéride tandis que les larves étaient la proie des moineaux, des grandes mésanges (Parus major) et des mésanges bleues (Parus caerulus). La grive (Turdus ericetorum) se nourrit des grosses larves de quatrième et cinquième instar. Les étourneaux ont été observés en train d'arracher les feuilles de chou pour aller chercher les larves au coeur des choux. Les plus importants prédateurs des chrysalides sont les oiseaux, particulièrement les mésanges. Moss (1933) évalue à environ 30% la réduction du nombre de chrysalides par les oiseaux. Au début du siècle, en Europe, il était conseillé de laisser aller des dindons dans les champs de crucifères, particulièrement tôt le matin, pour contrôler la piéride.
Autres prédateurs: Les araignées, les chrysopes, les coccinelles et les syrphides peuvent s'attaquer à tous les stades de l'insecte. Les libellules s'attaquent aux papillons. Il est possible de se procurer des chrysopes et de les relâcher au champ bien que cette méthode donne des résultats incertains. Quand les populations de ravageurs sont trop faibles, l'achat de chrysopes n'est pas justifié et quand les populations sont trop fortes, les chrysopes ne fournissent pas à la tâche.
Trichogrammes: Les trichogrammes sont des parasites qui pondent leurs oeufs dans les oeufs des piérides et autres lépidoptères. Le lâcher massif de trichogrammes dans les cultures est une technique employée depuis plusieurs années en Californie. François Fournier, d'Agriculture Canada à St-Jean-sur-Richelieu, étudie présentement l'optimisation de cette technique pour le Québec dans les cultures de crucifères. En pratique cependant, il est déjà possible de se procurer des trichogrammes et de les relâcher pour contrôler la piéride. Les trichogrammes sont disponibles commercialement chez les firmes spécialisées dans le contrôle biologique (Voir à ce sujet la liste des références utiles du Centre d'Agriculture Biologique de La Pocatière).
Guêpes braconides: Les bracones Apanteles rubecula et Apanteles glomeratus sont des parasites européens de la piéride maintenant établis au Canada et qui ont fait l'objet de nombreuses études. Dans ce qui fut la première expérience de lutte biologique classique, Riley, en 1883 aux États-Unis, avait importé la guêpe grégaire Apanteles glomeratus d'Angleterre qui par la suite s'est propagée un peu partout en Amérique. Ainsi, au début du siècle au Québec, DuPorte (1913) rapportait que ce parasite était aussi abondant que la piéride dans toute la province. Apanteles rubecula n'a été introduite dans l'est du Canada qu'au début des années 1970 mais a réussi à s'établir dans certaines régions (Corrigan, 1982). Certains biotypes d'A. rubecula survivent mieux à l'hiver que d'autres (Sengonca et Peters, 1993). Il est donc important de tenir compte de ce fait dans les lâchers de ce parasite.
La femelle de ces bracones attaque les chenilles des trois premiers instars (57% le premier instar). Les femelles d'A. glomeratus pondent 25 oeufs ou plus dans une seule chenille, attaquant un maximum de 2.5 chenilles par jour pendant un peu plus de deux semaines. Les femelles d'A. rubecula ne pondent qu'un seul oeuf par chenille car leurs larves sont plus grosses que pour A. glomeratus. A. rubecula est donc un agent de lutte biologique plus efficace d'un point de vue économique. Les larves du parasite émergent de la chenille mature alors qu'elle se prépare à faire cocon. Il n'y a pas de différences de développement entre les chenilles parasitées ou non. Celles-ci ont donc le temps de faire des dommages considérables même si elles meurent avant la nymphose.
En Nouvelle-Zélande, Moss (1933) évaluait à environ 30% la mortalité naturelle des chenilles dû à A. glomeratus et A. rubecula. Ce chiffre est évidemment très variable d'un lieu ou d'une année à l'autre. Au Canada, Kelleher et Hulme (1984) considèrent que l'efficacité d'A. glomeratus est excellente, mais que son action se fait trop tard dans la saison, d'où l'importance des lâchers tôt en saison. Les lâchers impliquent de se procurer des bracones commercialement, ce qui ne semble pas possible au Québec présentement.
Au Missouri, Parker et Pinnell (1972) ont obtenu des dommages moindres que le seuil de tolérance économique en relâchant 5000 adultes d'Apanteles rubecula par acre de culture. La clé de leur succès consistait à relâcher des larves de piérides en même temps que les parasites. Contrairement aux trichogrammes qui n'étaient efficace que par beau temps, les guêpes Apanteles avaient tendance à être efficace même par temps froid et pluvieux et à mesure que la saison avancait (pour la culture tardive).
Autres guêpes: Le tachine Phryxe vulgaris attaque les larves des derniers instars. Les ichneumones Mesochorus tachypus, Astomaspis nanus, A. submarginus, Anilastus ebeninus parasitent aussi les larves. Les chalcides Tetrastichus rapo et Pteromalus puparum attaquent la pupe. Ce dernier est très efficace mais trop tard en saison.
Virus: Les chenilles meurent naturellement en grand nombre à cause d'un baculovirus dit granulosé (GV). La mortalité naturelle dûe à ce virus peut atteindre jusqu'à 94% selon Harcourt (1963). Au début de l'infection, la chenille paraît plus lente et plus pâle. Puis elle tourne au jaune laiteux et se gonfle. Après la mort, le corps devient noir, se rompt et les tissus liquéfiés se répandent sur les feuilles et le sol. Dans les chenilles affectées sévèrement par le virus, la survie du parasite A. glomeratus est réduite (Levin et al., 1981). L'efficacité du virus dépend de la température et de la dose (Tatchell, 1981). Jaques (1970) conseille d'appliquer le virus au sol plutôt qu'au feuillage de façon à ce qu'il soit plus persistant et que sa survie soit assuré dans le même champ pour les années à venir.
| Jus de
piérides virulentes Le virus qui tue naturellement les larves de piérides atteindra un plus grand nombre de victimes si vous encouragez sa propagation. Vous pouvez faire votre propre pulvérisation virale en ramassant une dizaine de chenilles malades (blanches), en les passant au mélangeur avec de l'eau. Pulvériser sur le sol et les plants à protéger. Les larves atteintes meurent en quelques jours. |
Bt: Le Bt (Bacillus thurigiensis) est le recours microbien par excellence pour tous les types de chenilles. Après ingestion du Bt, les larves cessent de manger mais ne meurent qu'au bout de quatre à cinq jours. Le Bt n'est pas très contagieux ce qui fait que l'on doit le réappliquer. Il n'affecte pas les mammifères et les auxiliaires.
Le contrôle exercé sur la piéride est jugé de bon à excellent par Yepsen (1984), aussi bon ou mieux que les insecticides chimiques par Kelleher et Hulme (1984). Des expériences récentes en laboratoires et en champ indiquent toutefois que certains insectes développent une résistance progressive au bout de plusieurs générations. C'est le cas de la fausse-teigne en Floride par exemple. Il serait donc indiqué dans un programme de lutte contre la piéride de ne pas se fier seulement au Bt. Il est important de bien couvrir le feuillage car le Bt doit être ingéré par les chenilles pour agir. Suivre les recommandations des fabriquants pour l'utilisation du Bt.
Pierre Martel, d'Agriculture Canada à St-Jean-sur-Richelieu, a comparé les différentes marques de Bt disponibles au Canada sur les lépidoptères dans les cultures de crucifères. Le Thuricide serait plus efficace que le Dipel et que le Bactospéine selon ses recherches.
Un meilleur contrôle peut être obtenu en mélangeant Bt et le virus granulosé selon Jaques (1977).
Un grand nombre de plantes peuvent être employées pour repousser les papillons ou tuer les larves de la piéride. Étant donné l'importance économique de l'insecte, plusieurs chercheurs se sont penchés sur le sujet.
Les répulsifs végétaux consistent en des purins de plantes ou des extraits de plantes que l'on pulvérise sur le feuillage de la plante à protéger afin d'en masquer l'odeur et d'en imposer une autre qui repoussera le ravageur. Contre la piéride, les plantes les plus intéressantes sont la tomate, l'épinard, la tanaisie et les crucifères que la piéride évite. Les chercheurs qui ont étudié les répulsifs végétaux expriment leurs résultats par rapport à un indice de répulsion qui s'échelonne de 1 à 100.
Crucifères: Même si cela peut paraître incroyable, Renwick et Radke (1985) ont utilisé des crucifères avec succès pour repousser les piérides et les empêcher de pondre. En effet, la piéride lorsqu'elle perçoit l'odeur de tissus végétaux de crucifères endommagés, croit avoir affaire à une plante où une de ces larves ou une larve d'autres lépidoptères est en plein développement. Pour augmenter les chances de survie des oeufs qu'elle pondra, elle évite donc les plantes qui ont une forte odeur de chou endommagé. Les macérations de chou et les extraits à l'éther de chou avaient des valeurs d'indice de répulsion de 35 et 28. Parmis les autres crucifères hôtes des piérides, la lépidie (extraits à l'hexane) a donné une valeur de 63. Les extraits de bourse-à-pasteur (Capsella bursa-pastoris) et de vélar giroflée (Erysimum cheiranthoides), toutes deux des crucifères où les piérides ne vont pas pondre, ont reçus des indices de répulsion de 75 et 85 pour les extraits à l'eau.
Dimock et Renwick (1991) ont continué les recherches sur l'utilisation des extraits de vélar giroflée contre la ponte de la piéride. Ils ont étudié les extraits au butanol pour constater que l'effet répulsif était de courte durée en champ (environ trois jours à la première application) et avait tendance à diminuer après quelques applications. Comme les feuilles de crucifères sont très cireuses, la pluie déloge facilement tout produit pulvérisé.
Tanaisie: La tanaisie contient des substances qui préviennnent la ponte et l'alimentation de la larve chez de nombreuses espèces d'insectes, entre autres le doryphore de la pomme de terre. Hough-Goldstein et Hahn (1992) ont étudié en laboratoire et en champ l'effet contre la piéride d'extraits à l'eau de tanaisie appliqués sur des plants de choux. Il ressort de leurs expériences que les extraits de tanaisie agissent contre la ponte et l'alimentation des larves de piéride, l'effet étant moins apparent en champ qu'en laboratoire. L'application hebdomadaire d'extraits à l'eau de tanaisie sur des plants de choux à permis de réduire significativement les dommages à la récolte mais s'est avérée moins efficace que l'utilisation du Bt. Les auteurs rapportent des problèmes de phytotoxicité reliés à l'emploi d'un surfactant. L'efficacité de la tanaisie pourrait être améliorée par l'emploi d'un surfactant approprié et d'un solvant autre que l'eau.
| Extraits de
tanaisie (5%) - Prendre 50 g de feuilles de tanaisie fraîches. - Les broyer au mélangeur avec un litre d'eau. - Filtrer le mélange à travers deux épaisseurs de coton fromage. - Utiliser immédiatement. - Pulvériser à la transplantation et une fois par semaine par après. |
Autres plantes: Lundgren (1975) en Suède a étudié l'effet répulsif de plusieurs plantes rapportées comme répulsives dans la littérature scientifique ancienne et populaire contre les piérides, surtout la grande piéride P. brassicae. Selon ses travaux, les plantes les plus efficaces en ordre décroissant de répulsion étaient: les pousses fraîches de tomate (100.0); les feuilles de tomates broyées (90.9); les extraits à l'éther de sureau noir (Sambucus nigra) (83.0); l'huile d'oignon (60.9); les pousses de tomates congelées (45.8). Abivardi et Benz (1984) ont aussi constaté que les extraits de menthe poivrée, d'angélique et d'eucalyptus étaient très défavorables à l'alimentation des larves de piérides.
Les extraits à l'eau de soya, de tomate et d'épinard ont donné des indices de répulsions de 85, 79 et 94 respectivement dans les expériences de (Rendwick et Radke, 1985). On peut conclure que des pulvérisations d'extraits d'épinards à l'eau peuvent empêcher très efficacement (presque complétement) la ponte des piérides. Pour produire un purin répulsif de tomate, on laisse macérer des feuilles et des pousses de tomates (drageons) dans de l'eau tiède ou chaude pendant 2 à 5 heures. On pulvérise dès l'apparition des papillons. La concentration optimale n'est pas précisée dans la litérature.
Tabashnik (1987) a pu empêcher les piérides de pondre sur des choux en couvrant le feuillage de rutine ou de coumarine, des composés que l'on retrouve chez un grand nombre d'espèces végétales.
Yepsen (1984) propose une bouillie répulsive faite au mélangeur avec de la menthe, des tiges d'oignons vertes, des feuilles et des racines de raifort, des piments rouges forts, des grains de poivre et de l'eau. On ajoute du savon pure, on dilue et on verse sur chaque plant.
Roténone: La roténone est une substance extraite de plusieurs légumineuses tropicales, entre autres Derris elliptica et Derris trifoliata. Les inconvénients de la roténone sont son coût élevé, sa non-sélectivité et sa toxicité pour les poissons. Il ne faut donc pas l'appliquer près des cours d'eau. La piéride est plus sensible à la roténone que la fausse arpenteuse (Trichoplusia ni) selon Huckett (1936). Les doses à appliquer varient selon la concentration de roténone dans le produit et selon qu'il s'agit d'un poudrage ou d'une pulvérisation. L'application doit être faite à l'apparition des papillons et répétée tous les sept à quatorze jours selon les précipitations.
POUDRAGE: D'après les différentes études examinées, une dose de 25 à 30 kg/ha d'une poudre contenant 0,5 à 1,0 % de roténone serait appropriée.
PULVÉRISATION: Une poudre contenant 0,5 % de roténone est diluée à raison de 1:10000 et appliquée à 550-750 l/ha (Huckett, 1934). Pyenson (1941) a obtenu un aussi bon contrôle avec aussi peu que 0,12 kg de roténone par 100 litres d'eau en ajoutant un adjuvant de poudre de lait écremé à raison de 0,36 kg/100 litres.

Pyrèthre: Le pyrèthre est nettement moins efficace que la roténone contre la piéride et ne saurait être conseillée. Les plants de choux traités au pyrèthre ou avec un mélange de pyrèthre et d'ail étaient plus attaqués que les témoins dans les expériences de Endersby et al. (1992). Le pyrèthre aurait même un effet phytotoxique sur le chou par temps chaud selon ces auteurs.
Sabadille: La sabadille est efficace contre la piéride (Brunn et Allen, 1945). Une poudre à 10% est aussi efficace qu'une à 20%. Une pulvérisation de 1kg/100 l d'eau donne des résultats supérieurs à une pulvérisation de 0.5 kg/100 l.
Autres: Le nim et le quassia agissent contre la piéride. L'extrait aqueux de l'ail n'a pas d'effet mais une émulsion faite à partir de l'huile est mortelle à la piéride (Greenstock, 1970). Les autres plantes semi-tropicales ou tropicales suivantes ont également une action contre la piéride:
La compagnie Rohm and Haas commercialise aux États-Unis un produit appelé Mimic qui n'affecte que les larves de lépidoptères. Il s'agit en fait d'une hormone qui contrôle la mue, sauf qu'elle provoque l'apparition d'une nouvelle peau à l'intérieur de la vieille peau, ce qui cause la mort de l'insecte en quelques jours.
Certaines plantes produisent des substances en tout point identiques aux hormones responsables de la mue chez les insectes. Les substances isolées par Sato (1968) de diverses plantes asiatiques avaient une forte action sur la fausse-teigne des crucifères mais par sur la piéride.
Le savon insecticide et la chaux appliqués directement sur les larves vont tuer les jeunes larves. Ces substances peuvent être appliquées dans les quelques jours avant la récolte. Selon les expériences en champ d'Endersby et al. (1992), le savon insecticide a très peu d'efficacité contre la piéride et peut avoir un effet dépresseur sur les rendements de choux.
Le truc de jardiniers consistant à appliqué du lait suri au centre des plants n'a pas résisté à un test scientifique réalisé à l'Université de l'Illinois. Un autre truc conseillé par les manuels de jardinage biologique est de poudrer les plants avec 1/2 tasse de sel et une tasse de farine lorsque la rosée est sur les plants deux fois par semaine. Lorsque les chenilles avalent cette mixture, elles gonflent et meurent. On peut aussi appliquer ces deux substances en pulvérisation avec de l'eau. De la farine de seigle appliquée après une pluie va coller aux chenilles et les étouffer.
Les cendres de bois et la terre diatomée sont également conseillés parfois. Certaines formulations commerciales de terre diatomée sont homologuées aux USA pour usage contre la piéride.
On peut conclure de cette revue de la litérature sur la piéride les points suivants:
Abivardi, C et G. Benz. 1984. Tests with the extracts of 21 medicinal plants for antifeedant activity against larvae of Pieris brassicae. Mitteilungen der Sweizerischen Entomologischen Gesellschaft, 57:383-392.
Altieri, M.A. 1984. Patterns of insect diversity in monocultures and polycultures of Brussels sprouts. Protection Ecology, 6:227-232.
Andaloro, J.T., A.M. Shelton et C.J. Eckenrode. 1982. Seasonal abundance of Lepidopterous larvae in commercial cabbage fields. Environmental Entomology, 11:144-146.
Andow, D.A., A.G. Nicholson, H.C. Wien et H.R. Willson. 1986. Insect populations on cabbage grown with living mulches. Environmental Entomology, 15:293-299.
Baker, R.R. 1970. Bird predation as a selective pressure on the immature stages of the cabbage butterflies, Pieris rapae and Pieris brassicae. Journal of Zoology, 162:43-59.
Beirne, B.P. 1971. Pest insects of annual crop plants in Canada. I. Lepidoptera. II. Diptera. III. Coleoptera. Memoirs of the Entomological Society of Canada No. 78. The Entomological Society of Canada, Ottawa.
Bosio, C.F., K.D. McRea, J.K. Nitao et W.G. Abrahamson. 1990. Defense chemistry of Solidago altissima: effects on the generalist herbivore Trichoplusia ni (Lepidoptera: Noctuidae). Environmental Entomology, 19(3):465-468.
Brett, C.H. et M.J. Sullivan. 1974. The use of resistant varieties and other cultural practices for control of insects on crucifers in North Carolina. North Carolina Agricultural Experiment Station Bulletin No. 449. 31 pages.
Brunn, L.K. et T.C. Allen. 1945. Effectiveness of sabadilla in control of cabbage worms. Journal of Economic Entomology, 38(3):392.
Chiu, S.-F. 1950. Effectiveness of chinese insecticidal plants with reference to the comparative toxicity of botanical and synthetic insecticides. Journal of the Science of Food and Agriculture, 1:276-286.
Cook, A.J. 1881. Experiments with pyrethrum:safe remedies for cabbage worms and potato-beetles. The American Naturalist, 15:145-147.
Corbet, P.S. 1974. Habitat manipulation in the control of insects in Canada. Proceedings of the Tall Lumber Conference on Ecological Animal Control by Habitat Management, 5:141-171.
Corrigan, J.E. 1982. Cotesia (Apanteles) rubecula (Hymenoptera:Braconidae) recovered in Ottawa, Ontario ten years after its release. Proceedings of the Entomological Society of Ontario, 113:71.
Courter, J.W. et R. Randell. 1975. Planting herbs in garden vegetables to repel harmful insects. pages 51-56 In Horticulture at the Dixon Springs Agricultural Center. University of Illinois, Department of Horticulture.
Cromartie, W.J. 1981. The environmental control of insects using crop diversity. pages 223-251 In Pimentel, D. CRC Handbook of pest management in agriculture. Volume I. CRC Press, Boca Raton, Floride.
Cromartie, W.J. 1975a. The effect of stand size and vegetational background on the colonization of cruciferous plants by herbivorous insects. Journal of Applied Ecology, 12:517-533.
Cromartie, W.J. 1975b.Influence of habitat on colonization of collard plants by Pieris rapae. Environmental Entomology, 4:783-784.
Crepps, W.F. et L.E. Ehler. 1983. Influence of specific non-crop vegetation on the insect fauna of small-scale agroecosystems. UC Appropriate technology program, University of California, Berkeley. Research Leaflet Series #19-78-5.
Culliney, T.W. 1985. Predation of the imported cabbageworm Pieris rapae by the stink bug Euschistus servus euschistoides (Hemiptera:pentatomidae). The Canadian Entomologist, 117:641-642.
Dempster, J.P. 1969. Some effects of weed control on the numbers of the small cabbage white (Pieris rapae) on Brussels sprouts. Journal of Applied Ecology, 6:339-345.
Dempster, J.P. et T.H. Coaker. 1974. Diversification of crop ecosystems as a means of controlling pests. In Jones, D.P. et M.E. Solomon. Biology in pest and disease control. Blackwell Scientific Publications, Oxford, Royaume-Uni.
Dimock, M.B. et J.A.A. Renwick. 1991. Oviposition by field populations of Pieris rapae deterred by an extract of a wild crucifer. Environmental Entomology, 20(3):802-806.
Eigenbrode, S.D. et D. Pimentel. 1988. Effects of manure and chemical fertilizers on insect pest populations on collards. Agriculture, Ecosystems and Environment, 20:109-125.
Ellis, P.R., J. A. Hardman, M. Hommes, R. Dunne, S. Fischer, J. Freuler, A. Kahrer et C. Terretaz. 1988. An evaluation of supervised systems for applying insecticide treatments to control aphid and foliage caterpillar pests of cabbage. Brighton Crop Protection Conference.
Endersby, N.M., W.C. Morgan, B. C. Stevenson et C.T. Waters. 1992. Alternatives to regular insecticide applications for control of lepidopterous pests of Brassica oleracea var. capitata. Biological Agriculture and Horticulture, 8:189-203.
Endersby, N.M. et W.C. Morgan. 1991. Alternatives to synthetic chemical insecticides for use in crucifer crops. Biological Agriculture and Horticulture, 8:33-52.
Gossard, T.W. et R.E. Jones. 1977. The effects of age and weather on egg-laying in Pieris rapae. Journal of Applied Ecology, 14:65-71.
Gould, W.P et R.L. Jeanne. 1984. Polistes wasps (Hymenoptera: Vespidae) as control agents for lepidopterous cabbage pests. Environmental Entomology, 13:150-156.
Greenstock, D. 1970. Garlic as a pesticide. Henry Doubleday Research Association, UK.
Harcourt, D.G. 1962. Biology of cabbage caterpillars in Eastern Ontario. Proceedings of the Entomological Society of Ontario, 93:60-75.
Harcourt, D.G. et L.M. Cass. 1959. Control of caterpillars on cabbage in the Ottawa valley of Ontario and Quebec, 1956-1957. Journal of Economic Entomology, 52(2):221-223.
Harcourt, D.G., R.H. Backs et L.M. Cass. 1955. Abundance and relative importance of caterpillars attacking cabbage in Eastern Ontario. The Canadian Entomologist, 87:400-406.
Harrison, P.K. et R.W. Brubaker. 1943. The relative abundance of cabbage caterpillars on cole crops grown under similar conditions. Journal of Economic Entomology, 36(4):589-592.
Heath, R.R., P.J. Landolt, B. Dueben et B. Lenczewski. 1992. Identification of floral compounds of night-blooming jessamine attractive to cabbage looper moths. Environmental Entomology, 21(4):854-859.
Hough-Goldstein, J. et S.P. Hahn. 1992. Antifeedant and oviposition deterrent activity of an aqueous extract of Tanacetum vulgare on two cabbage pests. Environmental Entomology, 21(4):837-844.
Hovanitz, W. et V.C.S. Chang. 1964. Adult oviposition responses in Pieris rapae. Journal of Research on the Lepidoptera, 3(3):159-172.
Huang, X. et J.A.A. Renwick. 1993. Differential selection of host plants by two Pieris species: the role of oviposition stimulants and deterrents. Etomol. Exp. Appl., 68:59-69.
Huckett, H.C. 1936. Some tests of pyrethrum, derris and nicotine mixtures against cabbage worms. Journal of Economic Entomology, 29(3):575-580.
Huckett, H.C. 1934. Field tests on Long Island of derris as an insecticide for the control of cabbage worms. Journal of Economic Entomology, 27:440-445.
Ignoffo, C.M. et D.B. Boucias. 1992. Relative activity of geographical isolates of Nomuraea bioassayed against the cabbage looper and velvetbean caterpillar. Journal of Invertebrate Pathology, 59:215-217.
Jansson, R.K., G.L. Leibee, C.A. Sanchez et S.H. Lecrone. 1991. Effects of nitrogen and foliar biomass on population parameters of cabbage insects. Entomol. Expl. Appl., 61:7-16.
Jaques, R.P. et D.R. Laing. 1978. Efficacy of mixtures of Bacillus thurigiensis, viruses, and chlordimeform against insects on cabbage. Canadian Entomologist, 110:443-448.
Jaques, R.P. 1977. Field efficacy of viruses infectious to the cabbage looper and imported cabbageworm on late cabbage. Journal of Economic Entomology, 70(1):111-118.
Jones, R.E. 1977. Movement patterns and egg distribution in cabbage butterflies. Jouranl of ANimal Ecology, 46:195-212.
Jones, R.E. 1987. Ants, parasitoids and the cabbage butterfly Pieris rapae. Journal of Animal Ecology, 56:739-749.
Kelleher, J.S. et M.A. Hulme. 1984. Biological control programs against insects and weeds in Canada 1969-1980. CAB, Wallingford, UK.
Kirby, R.D. et J.E. Slosser. 1981. A sequential sampling plan for three lepidopterous pests of cabbage. The Southwestern Entomologist, 6(3):195-200.
Klijnstra, J.W. 1985. Interspecific egg load assessment of host plants by Pieris rapae butterflies. Entomol. Exp. Appl., 38:227-231.
Koehler, C.S., L.W. Barclay et T.M. Kretchun. 1983. Companion plants. California Agriculture, 37:11-12.
Landolt, P.J. 1993. Effects of host plant leaf damage on cabbage looper moth attraction and oviposition. Entomol. Exp. Appl., 67:79-85.
Landolt, P.J., B. Lenczewski et R.R. Heath. 1991. Lure and toxicant system for the cabbage looper (Lepidoptera:Noctuidae). Journal of Economic Entomology, 84(4):1344-1347.
Latheef, M.A. et R.D. Irwin. 1979. The effect of companiate planting on lepideptoran pests of cabbage. Canadian Entomologists, 111:863-864.
Létourneau, D. 1988. Nitrogen fertilizer's effects on pests and predators. The Cultivar, 6(2):5-6.
Levin, D.B., J.E. Laing et R.P. Jaques. 1981. Interactions between Apanteles glomeratus (Hymenoptera:Braconidae) and granulosis virus in Pieris rapae (Lepidoptera:Pieridae). Environmental Entomology, 10:65-68.
Lundgren, L. 1975. Natural plant chemicals acting as oviposition deterrents on cabbage butterflies (Pieris brassicae, P. rapae and P. napi). Zoologica Scripta, 4:253-258.
Maguire, L.A. 1984. Influence of surrounding plants on densities of Pieris rapae eggs and larvae on collards. Environmental Entomology, 13:464-468.
Metcalf, R.L. et R.A. Metcalf. 1993. Destructive and useful insects. McGraw-Hill, New York.
Moss, J.E. 1933. The natural control of the cabbage caterpillars, Pieris spp.. Journal of Animal Ecology, 2:210-231.
Myers, J.H. 1985. effect of physiological condition of the host plant on the ovipositional choice of the cabbage white butterfly, Pieris rapae. Journal of Animal Ecology, 54:193-204.
Nealis, V. 1985. Diapause and the seasonal ecology of the introduced parasite, Cotesia (Apanteles) rubecula(Hymenoptera:braconidae). The Canadian Entomologist, 117:333-342.
Neupane, F.P. et D.M. Morris. 1991. Alpha-tocepherol alteration of soybean antiherbivory to Trichoplusia ni larvae. Journal of Chemical Ecology, 17(10):1941-1951.
Nieuwhof, M. 1969. Cole crops: botany, cultivation and utilization. World crops books. Leonard Hill, Londres. 353 pages.
Parker, F.D. et R.E. Pinnell. 1972. Further studies of the biological control of Pieris rapae using supplemental host and parasite releases. Environmental Entomology, 1(2):150-157.
Pimentel, D. 1961. An evaluation of insect resistance in broccoli, brussels sprouts, cabbage, collards, and kale. Journal of Economic Entomology, 54: 156-158.
Pyenson, L. 1941. Concentrations of derris sprays effective on the imported cabbage worm. Journal of Economic Entomology, 34(3):473-474.
Radcliffe, E.B. et R.K. Chapman. 1965. Seasonal shift in the relative resistance to insect attack of eight commercial cabbage varieties. Annals of the Entomological Society of America, 58(6):892-902.
Radcliffe, E.B. et R.K. Chapman. 1966. Varietal resistance to insect attack in various cruciferous crops. Journal of Economic Entomology, 59(1):120-125.
Renwick, J.A.A. et C.D. Radke. 1988. Sensory cues in host selection for oviposition by the cabbage butterfly, Pieris rapae. Journal of Insect Physiology, 34(3):251-257.
Renwick, J.A.A. et C.D. Radke. 1985. Constituents of host- and non-host plants deterring oviposition by the cabbage butterfly, Pieris rapae. Entomol. Exp. Appl., 39:21-26.
Richards, O.W. 1940. The biology of the small white butterfly (Pieris rapae), with special reference to the factors controlling its abundance. Journal of Animal Ecology, 9:243-288.
Root, R.B. et P.M. Kareiva. 1984. The search for resources by cabbage butterflies (Pieris rapae): eocological consequences and adaptive significance of markovian movements in a patchy environment. Ecology, 65(1):147-165.
Samson, P.R. et P.W. Geier. 1983. Induction of crop damage by the cabbage white butterfly, Pieris rapae(Lepidoptera: pieridae), on cabbage. Protection Ecology, 5:199-223.
Sato, Y. 1968. Insecticidal action of phytoecdysones. Applied Entomology and Zoology, 3(4):155-162.
Schoonhoven, L.M., E.A.M. Beerling, R. Braaksma et Y. Van Vugt. 1990. Does the imported cabbageworm, Pieris rapae, use an oviposition deterring pheromone? Journal of Chemical Ecology, 16(5):1649-1655.
Sengonca, C. et G. Peters. 1993. Biology and effectiveness of Apanteles rubecula (Hym., Braconidae), a solitary larval parasitoid of Pieris rapae (Lep., Pieridae). Journal of Applied Entomology, 115:85-89.
Sievers, A.F., W.A. Archer, R.H. Moore et E.R. McGovran. 1949. Insecticidal tests of plants from tropical America. Journal of Economic Entomology, 42(3):549-551.
Slansky, F. et P. Feeny. 1977. Stabilization of the rate of nitrogen accumulation by larvae of the cabbage butterfly on wild and cultivated plants. Ecological Monographs, 47:209-228.
Smith, C.L. et Z. Smilowitz. 1976. Growth and development of Pieris rapae larvae parasitized by Apanteles glomeratus. Ent. Exp. Appl., 19:189-195.
Smith, J.G. 1976. Influence of crop background on aphids and other phytophagous insects on Brussels sprouts. Annals of Applied Biology, 83:1-13.
Srinivasan, K. et G.K. Veeresh. 1986. Economic analysis of a promising cultural practice in the control of moth pests of cabbage. Insect Sci. Applic., 7(4):559-563.
Stewart, J.G. et M.K. Sears. 1990. Effects of density of imported cabbageworm larvae on cauliflower yield and marketability. Proceedings of the Entomological Society of Ontario, 121:111-113.
Swingle, W.T. 1941. A chinese insecticidal plant, Tripterygium wilfordii, introduced into the United States. Science, 93:60-61.
Tabashnik, B.E. 1987. Plant secondary compounds as oviposition deterrents for cabbage butterfly, Pieris rapae (Lepidoptera:pieridae). Journal of Chemical Ecology, 13(2):309-316.
Tatchell, G.M. 1981. The effects of a granulosis virus infection and temperature on the food consumption of Pieris rapae (Lepidoptera: Pieridae). Entomophaga, 26:291-299.
Theunissen, J., H. den Ouden et A.K.H. Wit. 1985. Feeding capacity of caterpillars on cabbage, a factor in crop loss assessment. Entomol. Exp. Appl., 39:255-260.
Theunissen, J. et H. den Ouden. 1980. Effects of intercropping with Spergula arvensis on pests of Brussels sprouts. Ent. Exp. Appl., 27:260-268.
Traynier, R.M.M. et R.J.W. Truscott. 1991. Potent natural egg-laying stimulant for cabbage butterfly Pieris rapae. Journal of Chemical Ecology, 17(7):1371-1380.
Traynier, R.M.M. 1979. Long-term changes in the oviposition behaviour of the cabbage butterfly, Pieris rapae, induced by contact with plants. Physiological Entomology, 4:87-96.
Vail, K.M., L.T. Kok et T.J. McAvoy. 1991. Cultivar preferences of lepidopterous pests of broccoli. Crop Protection, 10:199-204.
Wang, M., Q. Hailin, K. Man et L. Yanzi. 1991. Insecticidal sesquiterpene polyol ester from Celastrus angulatus. Phytochemistry, 30(12):3931-3933.
Wolfson, J.L. 1980. Oviposition response of Pieris rapae to environmentally induced variation in Brassica nigra. Ent. Exp. Appl., 27:223-232.
Yepsen, R.B. 1984. The encyclopedia of natural insect and disease control. Rodale Press, Emmaus, Pennsylvanie.
Zhao, J.Z., G.S. Ayers, E.J. Grafius et F.W. Stehr. 1992. Effects of neighboring nectar-producing plants on populations of pest lepidoptera and their parastoids in broccoli plantings. The Great Lakes Entomologist, 25(4):253-259.
1. Voir Traynier et Truscott (1991) pour plus d'information sur la nature de ces composés.
© 1994 Projet pour une agriculture écologique. Tous droits réservés.
Info Request | Services | Become EAP Member | Site Map
Give us your comments about the EAP site
Ecological Agriculture Projects, McGill University (Macdonald
Campus)
Ste-Anne-de-Bellevue, QC, H9X 3V9 Canada
Telephone:
(514)-398-7771
Fax:
(514)-398-7621
Email: info@eap.mcgill.ca
To report problems or otherwise comment on the structure of this site, send mail to the Webmaster